• MC-F-002. Capítulo 2. Hominización y longevidad.
2.1 Introducción
El objetivo de este capítulo es introducir al estudiante-lector en las características antropobiológicas que han ido determinando, a lo largo del tiempo de los homínidos, un incremento de la longevidad. En este capítulo mostraremos cómo la especie humana, que desde sus remotos orígenes, hasta hace un centenar de años (comienzo del siglo XX), tenía una esperanza de vida que no superaba la cuarta década, ha sobrepasado en la actualidad las siete décadas, al menos en los países desarrollados en menos de un siglo (Figura 1).
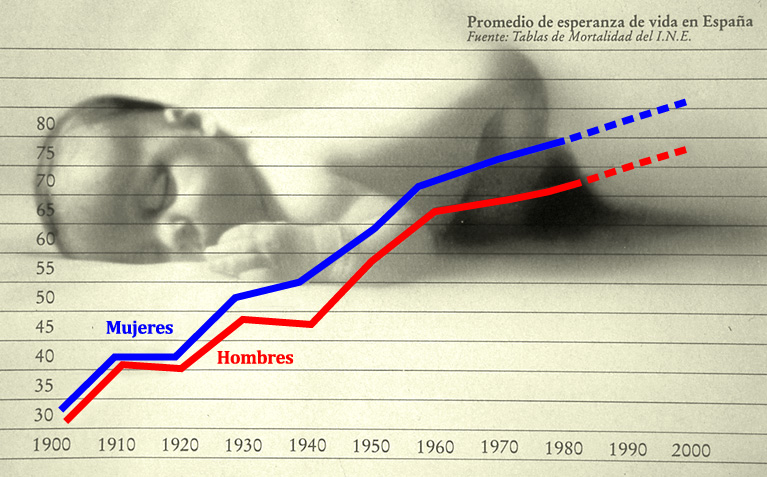
Figura 1. Tendencia del incremento de la esperanza de vida en España (adaptada de El País Semanal, 1980).
Como hemos visto, en el capítulo anterior, los seres vivos en el medio natural (hábitat salvaje), tienen una longevidad que puede ser incrementada al alojarlos bajo las condiciones controladas del laboratorio, zoológico, etc. Sin embargo ninguna otra especie se ha adaptado a los diferentes entornos como lo han hecho los humanos. Por medio de la culturización y la adaptación al medio, la humanidad ha organizado para ella un ambiente que la ha permitido expresar su potencial de longevidad. Esta adaptación ha tenido consecuencias, entre otras, sobre nuestro desarrollo y envejecimiento (Andrews y Pilbeam, 1996). En este sentido y con la idea inicial de tratar de explicar las causas del incremento de la longevidad humana analizaremos, inicialmente, la evolución de los homínidos, pero no es nuestro objetivo proponer una hipótesis de trabajo que ayude a resolver las incógnitas sobre el origen y evolución de nuestra especie. Sugerimos al lector interesado consultar bibliografía especializada en el tema, alguna de la cual es aquí citada.
Como punto de partida presentamos la tabla 1, en la cual se muestran las categorías y niveles taxonómicos de los humanos. El objetivo de esta tabla en insertar en el contexto taxonómico la situación del Homo sapiens sapiens, en comparación con otras especies próximas a nosotros (fundamentalmente los grandes simios actuales).
|
CATEGORÍAS Y NIVELES TAXONÓMICOS DE LA GENEALOGÍA HUMANA |
||
|
Reino |
Animalia |
Animales. |
|
Filo |
Chordata |
Animales con notocorda. |
|
Subfilo |
Vertebrata |
Animales con columna vertebral. |
|
Superclase |
Tetrapoda |
Animales con cuatro extremidades. |
|
Clase |
Mammalia |
Hembras con glándulas mamarias. |
|
Subclase |
Theria |
Mamíferos que conciben crías fetales. |
|
Infraclase |
Eutheria |
Mamíferos con placenta. |
|
Orden |
Primates |
Primates. |
|
Suborden |
Anthopoidea |
Todos los monos, simios y humanos. |
|
Superfamilia |
Hominoidea |
Simios y humanos. |
|
Familia |
Hominidae |
Los humanos y los antepasados inmediatos. |
|
Género |
Homo |
Miembros extintos y actuales de la especie. |
|
Especie |
Homo sapiens |
Especie humana moderna. |
|
Subespecie |
Homo sapiens sapiens |
Todos los seres humanos actuales. |
Tabla 1. Existen ciertas controversias entre los investigadores en la asignación de algunos de estos niveles. Por ejemplo hay antropólogos y biólogos que consideran que todos los humanos actuales pertenecen a la especie H. sapiens y no aceptan el término subespecie (elaboración propia).
2.2 Origen y evolución de los homínidos
En la actualidad y aunque existe gran controversia general sobre el origen de nuestra especie, parece bastante bien aceptado que los simios y los humanos hemos tenido unos ascendientes comunes, como han mostrado las pruebas para medir el grado de parentesco entre las especies vivas (Pilbeam, 2000). Analizando homologías morfológicas y bioquímicas de ADN de fósiles antiguos (ADNa) y las variaciones en aminoácidos en ciertas proteínas que las diferentes especies poseen en común (hemoglobina, insulina, etc.), se han podido establecer las relaciones inter-especies. Basándose en estos estudios se ha propuesto que los homínidos y los simios africanos han tenido un ancestro común hace 4-5 millones de años (crones).
La línea de los homínidos comenzó su diferenciación de los simios, probablemente, como consecuencia de iniciar un nuevo estilo de vida que comenzó a transcurrir, en su mayor parte, en el suelo y en un hábitat abierto (del tipo sabana). Este nuevo hábitat, alejado de la protección de las copas de los árboles, se cree determinó la aparición de aspectos relacionados con el desarrollo de la bipedestación y el caminar erguido. Además se inició el desarrollo anatómico del cerebro y los cambios conductuales asociados a dicho proceso. Estos cambios determinaron la aparición de las primeras adaptaciones anatómicas que separaron a los homínidos de los grandes simios. Entre estas primeras adaptaciones señalaremos como más características las que tuvieron lugar en la morfología de las extremidades inferiores (fundamentalmente los pies), las superiores (manos), el crecimiento del cráneo y la adaptación de la dentición a las nuevas fuentes de alimentos (Pilbeam, 1984).
|
ESTADIO EVOLUTIVO |
PERIODO EVOLUTIVO |
|
Australopithecus afarensis |
4-2.000.000 AÑOS |
|
Homo habilis |
2,5-1.000.000 AÑOS |
|
Homo erectus |
1,8-300.000 AÑOS |
|
Homo neanderthalensis |
500-35.000 AÑOS |
|
Homo sapiens arcaico |
300-20.000 AÑOS |
|
Homo sapiens sapiens |
35.000-ACTUALIDAD |
Tabla 2. Estadio evolutivo de la línea Homo y periodos temporales en los cuales existió cada una de la líneas que originaron el Homo sapiens sapiens (elaboración propia).
2.2.1 Los australopiteos (Australopithecus afarensis)
Los candidatos para el puesto de primeros antepasados (Tabla 2) separados ya de los simios corresponden a un grupo de fósiles encontrados en África datados entre 3,75-3,65 crones (Brunet et al., 1995). Estos restos óseos se atribuyeron y asignaron a un simio que ya presentaba algunas características que lo separaban de los otros simios coetáneos y lo aproximaban a la especie humana. Por esto se denominó Australopithecus afarensis, que significa “simio del sur”. Tres características importantes presentaban los A. afarensis que los incluían en la línea de la evolución humana: la bipedestación, la utilización de elementos naturales (ramas, piedras, etc.) como herramientas, y la disminución de la proyección de los dientes caninos al haber iniciado una alimentación basada, fundamentalmente, en la recolección de vegetales. Los australopitecos habitaban áreas de la actual Etiopía y la zona del Valle del Rift (Tanzania) en África. Los australopitecos, se ha sugerido, que habitaron estas zonas en un periodo comprendido entre los 4-2 millones de años y fue un grupo concreto de ellos el que dio un salto evolutivo al denominado Homo habilis. De esta manera debemos tener presente que los australopitecos y los habilis compartieron tiempo y espacio en la evolución (Figura 2). Los restos fósiles más famosos de un A. afarensis, debido a su abundancia y buen estado de conservación, pertenecen a una hembra que se denominó “Lucy” y que han sido datados con una antigüedad de 3,2 millones de años (Wolpoff, 1983).
2.2.1.1 Características morfológicas de los Australopithecus afarensis
Las características anatómicas más importantes de los australopitecos adultos, que se infieren del análisis de los fósiles encontrados son: la estructura de la pelvis y las extremidades sugiere que eran bípedos y tenían una capacidad de deambulación tipo humana. Esta anatomía adaptada al bipedismo ha sido considerada por los investigadores el rasgo fundamental para considerar a los afarensis como el inicio de la evolución de los homínidos (Wolpoff, 1983). El inicio de la marcha bípeda supuso un cambio en el centro de gravedad del cuerpo que produjo modificaciones en las curvaturas de la columna vertebral. La nueva distribución del peso corporal sobre los pies hizo que se desarrollase el arco plantar. La bipedestación también tuvo una repercusión fundamental sobre las extremidades superiores donde, en la mano, se desarrolló y perfeccionó la oposición del pulgar con los restantes dedos (base del inicio de las habilidades manuales). Los australopitecos machos más altos tenían una talla de unos 1,20 m, y pesaban alrededor de 50 kg, mientras que las hembras alcanzaban los 0,8 m, y pesan alrededor de 30 kg. Como se puede apreciar estas cifras hablan de un gran dimorfismo sexual en cuanto a valores antropométricos.
Los australopitecos poseían una facies prognata debido al gran prognatismo de la mandíbula y la presencia de un hueso frontal muy huidizo que determinaba la apariencia de una frente aplanada. Respecto al tamaño cerebral, los análisis de los volúmenes craneales sugieren que tenían una capacidad craneal con un rango de 350-500 cm3. Las capacidades más pequeñas corresponden a los australopitecos más antiguos y las más próximas a 500 cm3 corresponden a los A. afarensis más tardíos ya en transición hacia el primitivo Homo. Sirva como referencia que la media de la capacidad craneal de los chimpancés actuales tiene un rango de 300-485 cm3. Se ha sugerido que el progresivo incremento del volumen cerebral a lo largo de la evolución de los homínidos guarda una estrecha relación con los cambios en el esqueleto óseo, previamente comentados, y de forma particular con el desarrollo de las habilidades manuales fundamentadas en la oposición del pulgar que permitió a los australopitecos manejar objetos con cierta destreza y de esta manera actuar sobre las áreas corticales sensorio-motrices y determinar su desarrollo y maduración (Strait y Grine, 2004).
2.2.1.2 El ciclo vital de los Australopithecus afarensis
El australopiteco y el hombre moderno, comparten algunos rasgos con los primates antropoides, nuestros parientes evolutivos más cercanos. Desde el punto de vista de la longevidad nos interesa destacar: 1) embarazos prolongados (humanos 260-280 días, chimpancés 240-260, y gorilas 260-280 días), 2) una cría por parto y muy raramente partos múltiples, 3) un largo periodo de dependencia infantil, 4) intensa vida social y, 5) grandes cerebros en relación con la masa corporal (Bercovitch y Harding, 1993). Se ha sugerido, que al igual que sucede en la actualidad con los primates, los australopitecos tenían una relación de 4 hembras por cada varón que nacía. Esto determinaba, junto con la más precoz maduración sexual de las hembras, la posibilidad de que siempre hubiera más hembras disponibles para incrementar la capacidad reproductiva de la especie (darwinismo). Este es un rasgo que se ha perdido con la evolución humana donde, en la actualidad, la ratio de alumbramientos hembras/varones es casi 1.
El estudio de los fósiles de los Australopithecus ha revelado que presentaban un ciclo vital similar al de los simios coetáneos, particularmente en lo que se refiere a las primeras etapas del desarrollo postnatal. Ambos compartían un rápido desarrollo de las fases postnatales tempranas para alcanzar, las hembras, la etapa adulta a los 5-7 años, mientras que los machos la alcanzaban más tarde, en torno a los 8-10 años. Es decir, tenían un desarrollo físico postnatal de tipo precocial (Bogin, 1999). Se entiende por tipo precocial (precoz) un rápido crecimiento para que en pocos años pudieran alimentarse, defenderse, cooperar, alcanzar su autonomía funcional, y en última instancia comenzar la reproducción. Una vez alcanzada la etapa reproductiva, las hembras podían pasar por una serie de ciclos embarazo-parto-lactancia habiéndose calculado que este ciclo se podía repetir cuatro veces y, la hembra llegaba al final de su etapa fértil en torno a los 20 años. Alcanzada esta edad, tanto en machos como hembras, se iniciaba un rápido proceso de envejecimiento (edentulismo, pérdida de fuerza, etc.), que determinaba su muerte por inanición o bajo la acción de un predador. De esta forma la longevidad máxima de los australopitecos se ha establecido en 25 años. Pero debemos señalar que eran muy pocos los que alcanzaban esta edad, pues la mayoría moría joven como consecuencia de infecciones, traumatismos y acción de los predadores. La esperanza de vida de los A. afarensis debido a la alta tasa de mortalidad infantil no superaba los 10 años (Crews y Gerber, 2003).
2.2.2 Los Homo (habilis y erectus)
Los A. afarensis que evolucionaron hacia la línea Homo fueron desarrollando durante varios millones de años sus habilidades manuales y unos cuerpos más robustos, además de incrementar el tamaño del cráneo (Figura 2). De esta manera, hace 1,9 crones, se originó el denominado Homo habilis que ya constituyen la emergencia definitiva del género Homo (Leakey, 1996). El nombre de Homo habilis (hombre hábil) se le otorgó en reconocimiento de la presumible asociación entre el H. habilis y el inicio de la fabricación de algunos instrumentos de piedra. Homo habilis habitó las mismas zonas que los australopitecos y la etapa evolutiva de los habilis comprende un periodo de tiempo entre 2,5-1 millones de años. Lo cual nuevamente nos señala la coexistencia de grupos de australopitecos y hábiles.
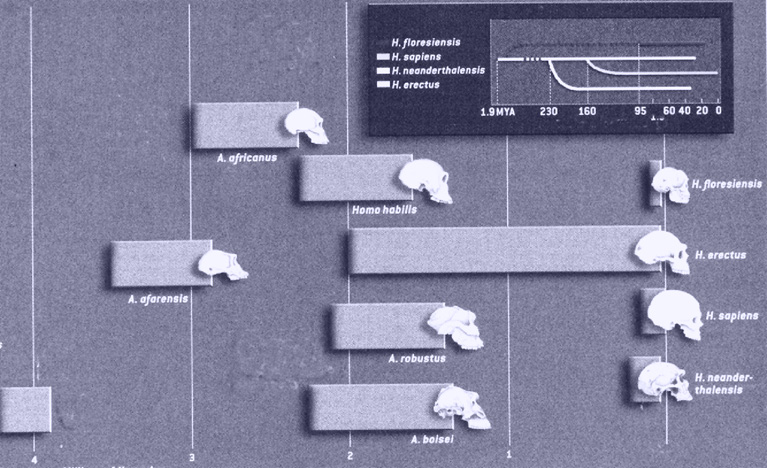
Figura 2. Posibles líneas de evolución de los homínidos. Observe la “discontinuidad” de esta evolución y la posición inicial de los Australopithecus afarensis. Se incluye el recientemente catalogado Homo floresiensis (adaptado de K. Wong, 2004).
2.2.2.1 Características morfológicas de los Homo habilis
Los habilis, como consecuencia de la estimulación de las áreas corticales motoras y sensitivas, iniciaron un proceso de incremento del talla, peso cerebral y, tal y como revelan los estudios de endovaciados poseían una capacidad craneal entre 500-750 cm3. La talla de las hembras era de unos 100 cm, y pesaban en torno a 35 kg y la de los varones alcanzaba los 140 cm y tenían un peso de poco más de 55 kg. Esto nos muestra el inicio de la longinización de nuestra especie. Además se ha propuesto que ya habilis era capaz de manejar el fuego que se ocasionaba de forma natural, aunque era incapaz de producirlo (Robinson, 1966).
Las características anatómicas más típicas de los habilis fueron: su progresiva estilización, el inicio del desarrollo de la bóveda craneal, la disminución del prognatismo y la reducción aún mayor que en la etapa anterior del tamaño de las piezas dentarias. Este aspecto de la reducción del tamaño de las piezas dentarias, fundamentalmente el tamaño de los caninos, determinó que su facies comenzara a tener rasgos del hombre moderno y tal vez más importante, justificó un tipo especial de alimentación que pasó a ser de tipo recolector (de alimentos estacionales) y los aportes de proteínas animales procedían fundamentalmente de pequeños animales cazados en grupo y de los restos de otros animales muertos. Se ha sugerido que este inicio de una alimentación de tipo omnívora dio un nuevo impulso al proceso de incremento del tamaño cerebral.
2.2.2.2 Ciclo vital de los Homo habilis
Respecto del ciclo vital de los habilis éste no presenta grande variaciones con respecto a lo señalado para los australopitecos. Se produce un pequeño incremento en la etapa reproductiva lo que permite a las hembras tener más descendencia. Estos hechos coinciden con lo que se denomina una selección-r. La selección-r es característica de los seres vivos que para alcanzar el éxito reproductivo y concretamente la propagación de esa especie, alcanzan de forma temprana la edad fértil, tienen altas tasas de reproducción (más descendencia), e invierten poco tiempo y esfuerzo en los cuidados a las crías. A esto se une el hecho de la cooperación social intra e inter grupos de habilis, lo que determina que disminuya la presión de los depredadores (acciones de defensa colectiva). Asimismo se cree que comenzaron a utilizarse remedios naturales para ciertas enfermedades (ingesta de ciertos vegetales). Esto determinó que aunque la longevidad máxima, con respecto a los australopitecos no se incrementó, si se produjo un aumento en la esperanza de vida que se ha calculado próxima a los 15 años (Wood, 1996).
2.2.3 Del Homo erectus al Homo sapiens arcaico (neanderthalis)
Los restos fósiles de H. habilis y los objetos encontrados en su entorno, sugieren que vivían en grupos o cuadrillas semejantes a los grupos de bandas de cazadores-recolectores de la actualidad y como resultado de, entre otras influencias, la progresiva organización social y el incremento de sus destrezas continuaron evolucionando y hace unos 1,8 millones de años se originó una especie llamada Homo erectus (Hombre erguido). Los erectus, en conjunto, vivieron desde la fecha señalada hasta hace unos 300.000 años (Antón, 2003).
2.2.3.1 Características morfológicas de los Homo erectus
Los primeros erectus fueron coetáneos de los últimos habilis, aunque parece que sólo fue un compartir tiempo evolutivo pero no de hábitat. H. erectus, durante su evolución sufrió transformaciones físicas importantes; se produjo un incremento de la talla por, fundamentalmente, el alargamiento de las extremidades inferiores. Además se hizo más longilíneo, pues al incremento en talla (160 cm para los varones y 130 cm para las hembras), le acompañó un discreto incremento de peso similar para ambos sexos, así los varones alcanzaban los 65 kg y las hembras alcanzaban los 45 kg (Crews y Gerber, 2003). A estas variaciones antropométricas se asociaban mejoras en la anatomía de la mano lo que a su vez seguía teniendo su influencia sobre el desarrollo cerebral, fundamentalmente el córtex. Estos cambios morfo-funcionales se tradujeron en un crecimiento de la capacidad craneana que albergaba un cerebro cada vez mayor y con el inicio de las circunvoluciones. El estudio de los ya numerosos restos fósiles de los erectus ha mostrado que poseía una capacidad craneal entre 800-1000 cm3, aunque su morfología seguía siendo de acusado prognatismo y las tablas de los huesos del cráneo eran muy gruesas. Plenamente adaptado al caminar erguido y dotado de un sistema músculo-esquelético robusto el erectus era fuerte y vigoroso.
El “erguido” desarrolló una gran capacidad de bipedestación y desplazamiento y, algunos grupos comenzaron a desplazarse de forma importante, desde su hábitat inicial, realizando la primera “salida de África” dirigiéndose hacia Eurasia (desde España a China e Indonesia). Estas migraciones supusieron un nuevo peldaño en la evolución de los homínidos, pues, representaron una nueva adaptación de esta especie del clima cálido en África a los climas más fríos de Eurasia. Se ha propuesto que esta adaptación al frío está en la base de dos aspectos fundamentales de la civilización: en primer lugar el dominio del uso del fuego para calentarse, defenderse y preparar alimentos y, en segundo lugar el inicio de la cultura asociada a la protección del frío ambiental (habitar cavernas y abrigarse con pieles). Los erguidos se asociaban en bandas de cazadores-recolectores donde ya había una clara distribución de las funciones sociales basadas en la dicotomía macho-hembra (los varones cazaban y las mujeres recolectaban). La cooperación necesaria para la caza, junto con las habilidades que su éxito implica, se ha propuesto como un nuevo impulso en el proceso de “cerebrización”, pues se cree que el desarrollo de la caza está en la base del inicio del lenguaje en los homínidos (protolenguaje).
2.2.3.2 El ciclo vital del Homo erectus
En el Homo erectus, se alargó la duración de las fases tempranas (infancia) lo que contribuyó a un crecimiento más lento hasta alcanzar el estado adulto. Es decir, se incrementó la duración de la etapa pre-reproductiva. De esta manera, lo que era un rápido tránsito entre infancia y estado adulto, pasó a tener un ciclo de tres fases (infancia-pubertad-adulto). Esto supuso un incremento en la duración del ciclo vital de varios años, pero las fases de adulto (reproductora) y envejecimiento se mantuvieron constantes en cuanto a su duración (Crews y Gerber, 2003). El tener más tiempo postnatal para alcanzar la maduración sexual (infancia-adolescencia), permitió a los H. erectus comenzaran a tener crías más inmaduras en cuanto al desarrollo de su soma y destinar los esfuerzos del desarrollo prenatal a la maduración del cerebro. Este aspecto en la mejora en la maduración cerebral intrauterina se ha considerado fundamental en el proceso de hominización. Así, el cerebro comenzó a incrementar sus circunvoluciones y de esta manera se produjo un importante aumento en la extensión de la superficie de este órgano (córtex cerebral). De esta manera se originó una dualidad en cuanto al estado de madurez de los H. erectus al nacer. Tenían un cerebro precocial (muy desarrollado y receptivo a la estimulación) y un soma altricial, menos maduro que necesitaba más atenciones postnatales.
A medida que se produjo una extensión en la etapa pre-reproductora para que el soma alcanzara su madurez, se incrementó el tiempo necesario para completar los procesos de la vida (crecimiento, desarrollo, maduración, reproducción y cría de los hijos). Esto originó un ligero aumento de la etapa post-reproductiva y en consecuencia un incremento de la longevidad, aunque la fase de envejecimiento seguía siendo breve. Los estudios predictivos y de análisis de fósiles establecen que estos H. erectus podían alcanzar los 25-30 años de vida (Crews y Gerber, 2003).
2.2.4 Del Homo erectus al Homo sapiens arcaico
El siguiente peldaño en la escalera de la evolución del Homo erectus tomó una bifurcación de extraordinaria importancia para la especie humana (Antón, 2003). Una línea evolucionó en las poblaciones que quedaron en África y otra en las que habían emigrado a Eurasia. Esta evolución originó en África los primeros Homo sapiens arcaicos hace unos 400-300.000 años. Estos sapiens arcaicos presentaban un cerebro más grande (una media de capacidad craneal de 1.200 cm3, aunque todavía tenían una frente pequeña y huesos de gran grosor. La “otra línea” de Homo erectus (la que colonizó Eurasia), siguió evolucionando para finalmente originar, hace aproximadamente 100.000 años, una subespecie de homínidos muy robusta, sin barbilla y frente muy inclinada, conocida como Homo sapiens neanderthalensis (Wood, 1996).
2.2.4.1 Características morfolígicas
La variedad Homo sapiens neanderthalensis (europea), tenía una apariencia robusta (talla de 170 cm para varones y 140 cm para las hembras, y un peso que superaba los 70 kg y los 60 kg respectivamente). Además, los neandertales tenían una capacidad craneal de 1.600 cm3 (superior a la media de los humanos actuales). Pero este incremento en la capacidad craneal no se correlacionaba con un incremento en la superficie del córtex cerebral, que seguía siendo pobre en circunvoluciones, como muestran los análisis de las huellas cerebrales en los huesos de la bóveda craneal. Se cree que muchos de los rasgos anatómicos de los neandertales representaban los efectos de la selección para la vida en climas extremadamente fríos (glaciaciones). Los restos encontrados han revelado que vivían en abrigos naturales que adaptaban y eran grandes cazadores. Una característica cultural importante de los neandertales es que comenzaron a enterrar a los muertos e incluir algo de su ajuar en el enterramiento.
2.2.4.2 Ciclo vital de los neandertales
Homo sapiens neanderthalensis vivió hasta hace unos 35.000 años, tiempo en el que desapareció constituyendo las causas de su eliminación uno de los enigmas más importantes en los orígenes del hombre actual. Entre las diversas hipótesis que se han propuesto para justificar su desaparición figura la dificultad que tenían las hembras para parir fetos de gran tamaño y cráneos muy osificados (sus fetos experimentaban un crecimiento corporal precocial) que dificultaba, sobremanera, los mecanismos del parto muriendo la madre y el hijo.
2.2.5 Del Homo sapiens arcaico al Homo sapiens moderno
Mientras, como hemos visto, el neandertal europeo se adaptaba a las condiciones frías de Europa y a su fatídica desaparición, sus contemporáneos en África (los H. erectus), que vivían en un ambiente cálido, se hacían aún más longilíneos y comenzaron a evolucionar en la dirección del Homo sapiens moderno cuyos primeros miembros aparecieron hace 100-70.000 años.
2.2.5.1 Características morfológicas del Homo sapiens arcaico
Esta evolución implicó entre otras características la reducción de las diferencias antropométricas morfo-sexuales con respecto a la talla y el peso (varones 170 cm, 65 kg y hembras 150 cm, 50 kg). Como se puede apreciar, con la evolución de los homínidos se va produciendo una paulatina reducción del dimorfismo sexual gracias a un incremento de los valores antropométricos de las hembras.
Los Homo sapiens arcaicos se caracterizaron por seguir incrementando la duración temporal de las etapas postnatales tempranas (pre-reproductiva) hasta la madurez sexual. Esto ha hecho que los humanos mostremos una fase fetal final (tres últimos meses) destinados fundamentalmente al desarrollo del cerebro y otras estructuras neurales en detrimento del desarrollo somático. Además se calcula que durante los 5 primeros años de la vida postnatal, se produce un desarrollo diferencial entre las partes neurales y las somáticas de nuestro organismo, de tal manera que al finalizar este periodo el sistema nervioso ha alcanzado un desarrollo morfológico que asegura el posterior desarrollo funcional.
2.2.5.2 El ciclo vital del Homo sapiens arcaico
Respecto a su longevidad se ha postulado que la longevidad máxima de esta especie no superaba los 40 años, aunque la esperanza de vida se alargaba al irse produciendo una mejora en las condiciones de vida (reducción de la mortalidad infantil, mejora e incremento en la diversidad de nutrientes aportados por la alimentación, existencia de “curanderos” versados en el arte de sanar, cooperación social, etc.). Estos aspectos, o como consecuencia de ello, fueron determinando que la relación entre hembras y varones al nacer se acercara a 2. De esta manera la poligamia necesaria en las etapas anteriores (4 hembras por varón) para seguir perpetuando la especie, ahora ya se veía reducida de forma notable y paulatinamente nos vamos acercando a grupos monogámicos, pues la tendencia una-uno (hembra-varón) se siguió continuando hasta las cifras actuales. Esta reducción en la relación hembras/varones determinó la paulatina aparición de grupos monogámicos, lo que determinó la selección de los cuidados a un solo miembro de la pareja por parte de los varones (aprovisionamiento, atenciones, etc.) lo cual se cree que socialmente determinó un incremento de la esperanza de vida de los sapiens (Crews y Gerber, 2003).
2.2.6 El Homo sapiens moderno
Según la hipótesis “Salir de África”, los Homo sapiens modernos evolucionaron a partir de algunos Homo sapiens arcaicos que se originaron en África (130-120.000 años) y hace unos 90.000 iniciaron la segunda emigración de África, ésta llena de éxito como lo muestran los resultados, que colonizó Asia (68.000 años), Australia (50.000 años), Europa (36.000 años) y América (15.000 años). De particular importancia es la colonización de Australia, pues se sabe que esta isla-continente nunca estuvo unida a Asia, lo cual infiere un gran desarrollo intelectual de los sapiens pues hizo falta la construcción de “balsas” y ciertos conocimientos de navegación de un número importante de elementos que aseguraran la procreación y reproducción de la especie. Esta hipótesis de “Salir de África” que como hemos visto postula un origen común, de los humanos actuales, de un ancestro africano se ha visto reforzada al aplicarse las nuevas técnicas de análisis del ADN mitocondrial (ADN mt), que se transmite de forma matrilineal. Estos estudios han mostrado que todos los miembros contemporáneos de la humanidad hemos tenido un antepasado común en África (Watson et al., 1997).
En Europa, los primeros sapiens modernos llegados de África, se denominaron Cromañón y, fueron coetáneos de los últimos Neandertales, aunque, tal y como muestran los estudios genéticos (ADN mt), no hubo “cruces” entre ambas poblaciones (Currat y Excoffier, 2004). Otra hipótesis que trata de explicar la desaparición de los Neandertales es la lucha por la supervivencia por recursos limitados con los “recién” llegados Cromañones, más inteligentes, hábiles y con mejor capacidad de adaptación a las variaciones del medio.
2.2.6.1 Características morfológicas del Homo sapiens moderno
No existen diferencias importantes en las características anatómicas entre los Homo sapiens modernos (Cromañones) y los hombres actuales, pero se ha propuesto que todos los humanos actuales pertenecemos a la subespecie Homo sapiens sapiens (Wood, 1996). Esta subespecie está integrada por muchas poblaciones con capacidad reproductora cruzada entre ellas, algunas de las cuales han sido tradicionalmente clasificadas como razas que han seguido procesos micro-evolutivos locales originando cuatro variedades: caucásico (leucodermo), negroide (melanodermo), mongólido (xantodermo) y una población más aislada de aborígenes australianos (austrálidos).
2.2.6.2 Ciclo vital del Homo sapiens moderno
El Homo sapiens moderno, hasta el advenimiento de la civilización (Homo sapiens sapiens), se caracterizó básicamente, por un inicio tardío de su reproducción, un gran esfuerzo en la cría de los hijos, engendrando descendientes de forma consecutiva en un periodo fértil de 10-30 años. Finalizada esta etapa se producía un rápido envejecimiento teniendo una longevidad máxima de unos 40 años. Aunque se cree que eran muy pocos los individuos que alcanzaban esta cifra (Crews y Gerber, 2003).
En la figura 3 se muestra de forma esquemática el proceso diferencial de maduración del sistema nervioso, concretamente el cerebro (inferido por el tamaño del cráneo) frente al soma (Crespo-Santiago, 2004). En los humanos actuales, los recién nacidos poseen un sistema nervioso muy desarrollado (precocial) y un soma muy inmaduro (altricial). En las etapas iniciales de la hominización (australopitecos y hábiles), los individuos nacían con un soma muy precocial y un sistema nervioso altricial. La necesidad de emplear más tiempo postnatal para alcanzar la maduración somática e iniciar la etapa reproductora ha tenido como consecuencia un incremento de la longevidad.
A modo de resumen de lo expuesto hasta aquí debemos señalar que aunque es muy tentador seguir el rastro de los diversos homínidos que han formado parte de nuestra línea evolutiva tratando de postular una teoría unitaria que pueda explicar la longevidad de la especie, nada más alejado de la realidad científica. Todas las hipótesis propuestas para explicar los orígenes de la humanidad, se centran en aspectos más o menos parciales y concretos. Por esta razón el proceso de intentar entender el origen y evolución de la especie humana quedaría totalmente incompleto si no se incluyera el aporte que el ambiente, en forma de cultura, ha realizado sobre los hábitos de vida para producir unos humanos cada vez más altriciales y longevos, desde el punto de vista físico, pero muy precociales desde el punto de vista neurológico (Figura 3). Esto incluye el incremento de la encefalización que permitió el perfeccionamiento del lenguaje, el control parcial sobre el medio ambiente, el advenimiento de la cultura, y la llegada de las estructuras sociales que han ido incrementando el desarrollo inicial del bienestar común.
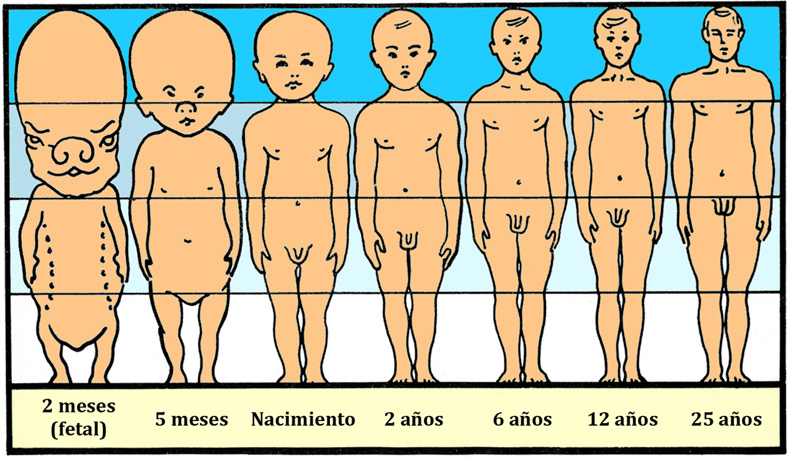
Figura 3. Muestra el tamaño relativo de la cabeza (cerebro) con respecto al soma desde el recién nacido al adulto. Se puede observar como en el momento del nacimiento la cabeza es 1/4 del tamaño corporal, mientras que en el adulto viene a representar 1/8. Así se entiende que los humanos actuales somos seres precociales en lo que a desarrollo cerebral se refiere, pero altriciales en el desarrollo físico (autor desconocido).
El punto de partida en el incremento decisivo de la longevidad humana se ha relacionado con el aumento temporal de la etapa de envejecimiento. Desde un punto de vista de la civilización esto se ha asociado con la llegada de la agricultura en las primeras culturas. Hasta esa fecha, tal y como hemos comentado (Tabla 3), la fase de envejecimiento era muy breve y los hombres no lograban sobrevivir más que unos pocos años al finalizar la etapa reproductora.
|
TIEMPO HISTÓRICO |
ESPERANZA DE VIDA |
LONGEVIDAD |
|
• Australopithecus afarensis • Homo habilis • Homo erectus • Homo neanderthalensis • Homo sapiens arcaico • Homo sapiens sapiens - PALEOLÍTICO SUPERIOR - MESOLÍTICO - NEOLÍTICO - EDAD DE BRONCE - EDAD DE HIERRO - IMPERIO ROMANO - ALTA EDAD MEDIA - BAJA EDAD MEDIA-XIV - SIGLO XV - SIGLOS XVI-XVII - XVIII - XIX - PRINCIPIOS DEL XX (*) - PRINCIPIOS DEL XXI |
10 20 20 20 25 25 28 30 29 22 23 20 26 20 30 33 34 40 40 80 |
25 25 30 30 35 40 40 – – – – – – – – – – – – 122 |
Tabla 3. (*) Hasta principios del siglo XX la esperanza de vida era la mitad de la actual. Es decir, en los últimos 100 años se ha doblado esta cifra. Asimismo, el Homo sapiens sapiens desde el Paleolítico Superior hasta principios del siglo XX sólo incrementó la esperanza de vida en 10 años. Hasta principios del siglo XX la esperanza de vida de las mujeres era inferior a la de los varones, pues había una alta mortalidad por partos. Desde esta fecha, la esperanza de vida de las mujeres ha superado a la de los varones en unos 5 años. Fuente. José Antonio Laceras (Director). Guía Artística Electra. Museo de Altamira pg. 38. MECyT. Barcelona
2.3 El envejecimiento desde el inicio de la civilización hasta comienzos del siglo XX
Los registros históricos muestran que en todas las etapas culturales de la humanidad (mesopotámica, egipcia, griega, romana, etc.) hasta el principio del XX, ha habido personas de una longevidad extrema para sus tiempo históricos (Tabla 4). Es fácil encontrar entre los grupos cultos, ricos o poderosos, de todas las culturas, personas que alcanzaron o superaron los 100 años, si bien su número es muy reducido. Lo más interesante es que la esperanza de vida de las diversas poblaciones se encontraba en torno a los 35 años. La mayoría de la gente moría a edades muy tempranas (mortalidad perinatal elevada). Desde un punto de vista de la reproducción, la gran mortalidad femenina impedía a muchas mujeres alcanzar la edad de la menopausia (Crews y Gerber, 2003). En Europa se ha calculado que al inicio del siglo XX sólo el 40% de las mujeres superaban la edad de 30 años, el 30% los 40 años y el 20% los 50 años (ver capítulos de biodemografía y transición epidemiológica).
|
HOMBRES Y MUJERES LONGEVAS |
||
|
Nacor (patriaca hebreo) |
2155-2008 a.C. |
(147 años) |
|
María (hermana de Moisés) |
1579-1452 a.C. |
(127 años) |
|
Adad-guppi (mujer noble de Babilonia) |
649-547 a.C. |
(102 años) |
|
Gorgias (filósofo sofista) |
483-378 a.C. |
(105 años) |
|
Hipócrates (“padre” de la medicina) |
460-365 a.C. |
(95 años) |
|
Cleantes (filósofo estoico) |
331-231 a.C. |
(100 años) |
|
Kai Wa (emperador japonés) |
213-98 a.C. |
(115 años) |
|
San Pablo de Tebas (ermitaño) |
229-341 |
(112 años) |
|
San Macario (obispo) |
300-390 |
(90 años) |
|
Acacio (obispo de Perea –Siria–) |
322-432 |
(110 años) |
|
Shenute (monje cristiano en Egipto) |
360-465 |
(105 años) |
|
San Millán (ermitaño) |
473-574 |
(101 años) |
|
Albumazar (astrónomo árabe) |
776-885 |
(109 años) |
|
Manegold de Lautenbach (filósofo alsaciano) |
1000-1103 |
(103 años) |
|
Simon Stock, San (religioso inglés) |
1165-1265 |
(100 años) |
|
Saadi (poeta persa) |
1195-1296 |
(101 años) |
|
García de Santa María, Alvar (escritor) |
1349-1460 |
(101 años) |
|
Columbela –Diveke– (noble de Dinamarca) |
1417-1517 |
(100 años) |
|
Anchieta, Miguel (escultor) |
1450-1588 |
(138 años) |
|
Silva, Feliciano de (escritor español) |
1451-1554 |
(103 años) |
|
Zamberti, Bartolomé (escritor italiano) |
1460-1560 |
(100 años) |
|
Nanek (profeta skiks) |
1469-1599 |
(130 años) |
|
Malinche (compañera de Hernán Cortés) |
1498-1597 |
(99 años) |
|
Agulló, Francisco (pintor) |
1538-1648 |
(110 años) |
|
Chifflet, Juan Jacobo (médico francés) |
1558-1660 |
(102 años) |
|
Antonio de Toledo (marqués de Mancera) |
1608-1715 |
(107 años) |
|
Legrand, Pedro (pirata francés) |
1632-1770 |
(138 años) |
|
Zapata, Diego Mateo (médico español) |
1644-1745 |
(101 años) |
|
Shaw, Meter (médico inglés) |
1695-1802 |
(107 años) |
|
Martín, Manuel José (impresor) |
1700-1800 |
(100 años) |
|
Sakai Hoitsu (poeta japonés) |
1721-1829 |
(127 años) |
|
Gómez, Ildefonso (militar) |
1731-1849 |
(118 años) |
|
Maturana y Vázques, Vicente (escritor) |
1739-1859 |
(120 años) |
|
Bere, Osvaldo (médico alemán) |
1742-1867 |
(125 años) |
|
Waldeck, Federico (artista checo) |
1766-1875 |
(109 años) |
|
González Peña, Carlos (periodista mejicano) |
1855-1955 |
(100 años) |
|
García, Manuel Vicente (barítono) |
1803-1906 |
(103 años) |
|
Robertson, Anna Mary (pintora EEUU) |
1860-1961 |
(101 años) |
|
Chiesa, Francesco (escritor suizo) |
1871-1973 |
(102 años) |
|
Sheldon, Wilmon Henry (filósofo EEUU) |
1875-1981 |
(106 años) |
|
Bayar, Celal (político turco) |
1884-1986 |
(102 años) |
|
Abbott, George (director teatral de EEUU) |
1887-1995 |
(108 años) |
|
Baline, Israel (compositor EEUU) |
1888-1989 |
(101 años) |
|
Mayr Ernst (evolucionista) |
1904-2005 |
(101 años) |
Tabla 4. Elaboración propia basada en la Enciclopedia Micronet 2000 y la Enciclopedia “El País” 2004. Tenga presente el lector que estos datos, fundamentalmente los referidos a las personas más antiguas no pueden resistir la crítica científica de los valores asignados a su ciclo vital. Hasta el Concilio de Trento (1545-1563) no se conocían registros parroquiales de natalicios y defunciones. En España el Registro se inició en 1870 (Ley Provisional del Registro Civil) que ha sufrido modificaciones posteriores, hasta las introducidas por el desarrollo de la Constitución Española de 1978 (elaboración propia).
La baja esperanza de vida de la humanidad hasta principios del siglo XX se debía fundamentalmente al azote de numerosas patologías que en la actualidad están superadas. Entre las importantes debemos señalar; la alta mortalidad perinatal por distocias, partos prematuros, incluyendo la mortalidad materna que hace que la esperanza de vida se vea notablemente reducida por eliminación muy temprana de personas en sus primeras etapas del ciclo vital. En segundo lugar señalaremos las enfermedades infecto-contagiosas que diezmaban de forma notable a la población al incidir en todos los niveles de las sociedades, aunque como es fácil de entender siempre las clases más desfavorecidas se veían más afectadas. Pandemias de peste, gripe, etc., afectaron de forma importante a todas las civilizaciones del mundo a lo largo de las etapas históricas. En tercer lugar, una gran parte de la población moría como consecuencia fundamental de la falta de higiene tanto personal como social. En este punto afectaban diarreas, alteraciones gastro-intestinales, etc. La introducción de sistemas de abastecimiento público de aguas controladas, el alcantarillado, recogida de basuras, etc., han permitido incrementar la longevidad gracias a medidas higiénicas preventivas. Finalmente, aunque en menor grado, se producían fallecimientos por fallos vasculares que afectaban al corazón y cerebro. A estas causas se añadían accidentes de todo tipo (laborales, guerras, etc.) que hacían que la población tuviera una esperanza de vida que no superaba los 40 años (Tabla 5).
El incremento de la mortalidad por causas degenerativas coincide con el incremento de la esperanza de vida. En la actualidad las enfermedades infecto-contagiosas juegan un pequeño papel en la mortalidad general pues las vacunaciones en la infancia y las campañas de prevención hacen que su incidencia sea baja. Por el contrario el alargamiento del ciclo vital, fundamentalmente unido a los procesos de desgaste corporal, han hecho que las enfermedades degenerativas estén ocupando un lugar destacado en el ranking de causas de muerte. Asimismo, y como hemos visto al estudiar la teoría inmunológica del envejecimiento, la disminución de las capacidades defensivas del organismo por reducción de las respuestas inmunes acrecienta la incidencia de cáncer por la reducción en la capacidad de nuestro sistema inmune en el reconocimiento y eliminación de las células con mutaciones (Vaupel, et al., 1998, Oeppen y Vaupel, 2002).
|
CAUSAS DE MUERTE |
|
|
HASTA PRINCIPIOS DEL SIGLO XX |
POSTERIOR AL SIGLO XX |
|
Neumonía y Gripe Diarrea y Enfermedades digestivas Tuberculosis Enfermedades cardiacas Enfermedades renales Accidentes (laborales) Cáncer Muerte perinatal (parto-puerperio) Malnutrición |
Enfermedades cardiacas Cáncer Lesiones cerebrales Enfermedades pulmonares Accidentes (circulación) Diabetes Enfermedades renales Infecciones (SIDA) Sociopatías |
Tabla 5. Se muestran las principales causas de muerte hasta comienzos del siglo XX y en la actualidad. Se puede observar que ahora las principales causas de mortalidad se relacionan con el incremento de la esperaza de vida, es decir, las enfermedades degenerativas (cerebrales, cardiacas, sistémicas) han tomado una gran preponderancia (elaboración propia).
2.4 El envejecimiento humano en la actualidad
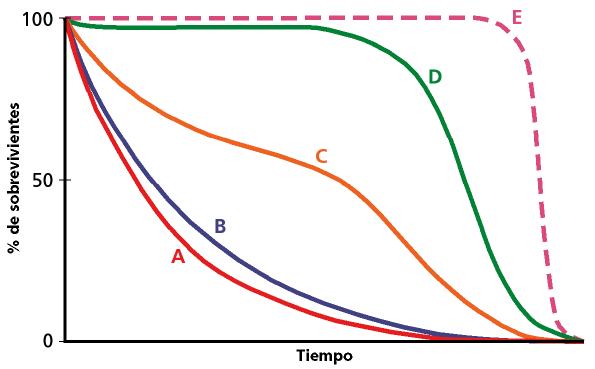
Figura 4. Nos muestra en forma de curvas (A, B, C, D, y E) diversas fases en la mortalidad de una especie determinada, en este caso la humana, afectada por el proceso de culturización y el avance biomédico (elaboración propia).
El gran incremento (casi un 100%) en la extensión del ciclo vital –longevidad–, está muy relacionada con el incremento de la duración del proceso de envejecimiento. Este aspecto de puede observar en la figura 4.
En el eje de abscisas se representa la hipotética duración de la vida en tiempo y en el de ordenadas el número de individuos. La curva A representa la mortalidad exponencial de una determinada población. En B se representa una población actual de cazadores-recolectores donde la mayoría de los individuos no superan los 30 años. En C se representa una población en países en vías de desarrollo, donde ya un mayor número de individuos alcanzan edades superiores a los 50 años. En D se representa la curva actual en los países desarrollados, en los cuales la mayor parte de los individuos alcanzan los 80 años. La curva E representa la dinámica “ideal” de una población, en la cual todos los individuos alcanzan la máxima longevidad. En todas las poblaciones siempre hay individuos que alcanzan la máxima longevidad.
Los humanos mostramos un desarrollo fetal en los tres últimos meses, destinados, fundamentalmente, al desarrollo del sistema nervioso y tres estadios pre-reproductivos (infancia, juventud y adolescencia). Durante la infancia se continúa el desarrollo y maduración del cerebro y los tejidos neurales asociados. Este desarrollo es muy rápido, alcanzando el 85% del peso y del tamaño que tendrá en el adulto, y se completa en el séptimo año. La fase juvenil está marcada por bajas tasas de crecimiento somático que precede a la adolescencia (edad entre 14-18 años). Durante esta última fase la dentadura madura, y las características físicas, fisiológicas y cognitivas se aproximan al adulto. El soma alcanza el desarrollo final en talla y los órganos reproductores maduran definitivamente.
El alargamiento del periodo de crecimiento y desarrollo del organismo es una estrategia evolutiva sólo válida para asegurar una baja mortalidad debida a peligros ambientales. Durante la edad reproductora se establece un tiempo suficiente para completar las tareas asociadas a la reproducción y a los cuidados parentales. Un modo sencillo de retardar (enlentecer) el desarrollo de un organismo complejo es la disminución del ritmo de la regulación endocrina, de la actividad del ADN, y de la síntesis de proteínas y respuestas celulares a dichas hormonas. La alteración hormonal de las fases del desarrollo altera fácilmente tanto el ritmo de crecimiento como la longitud del desarrollo, la maduración y el esfuerzo reproductivo.
Nunca antes en la historia de la humanidad habían tenido la mayoría de los humanos nacidos la oportunidad de vivir más allá de la cuarta década de vida. En la actualidad, al menos en los países desarrollados, la esperanza de vida se aproxima a los 80 años y parece que la tendencia es a un incremento progresivo de esta cifra. En este sentido la persona (bien datada) que más a vivido ha sido una mujer (Jeanne Calment, 1880-2002) que vivió 122 años y que aparece en la figura 5.
Como consideración final debemos señalar que la esperanza de vida, es un parámetro que puede ser afectado por diversas causas tanto intrínsecas como extrínsecas. Algunos países que poseían altas esperanzas de vida, debido a cambios político-económicos han visto reducido este parámetro en más de 20 años en sólo un periodo de 10 años, al entrar en fuertes crisis económicas que limitaron el destino de recursos socio-sanitarios.

Figura 5. Jeanne Calment, una mujer francesa que vivió 122 años. Falleció en 1997 pocos meses después de celebrar su último cumpleaños, aspecto que recoge esta foto. Ha sido la persona, bien datada, más longeva (autor desconocido).
Bibliografía
- Andrews P., Pilbeam D. Palaeoanthropology. The nature of the evidence. Nature. 1996;379:123-124.
- Anton S. C. Natural history of Homo erectus. Am. J. Phys. Anthropol. 2003;37:126-170.
- Bercovitch F. B, Harding R. S. Annual birth patterns of savanna baboons (Papio cynocephalus anubis) over a ten-year period at Gilgil, Kenya. Folia Primatol (Basel). 1993; 61:115-122.
- Bogin B. Evolutionary perspective on human growth. Annu. Rev. Anthropol. 1999;28:109-153.
- Brunet M., Beauvilain A., Coppens Y., Heintz E., Moutaye A. H., Pilbeam D. The first australopithecine 2,500 kilometres west of the Rift Valley (Chad). Nature. 1995;378:273-275.
- Crespo Santiago D. De la neurona al cerebro: la base de los procesos cognitivos. Pedrueca. 2004;1:10-13.
- Crews D. E., Gerber L. M. Reconstructing life history of hominids and humans. Coll. Antropol. 2003;27:7-22.
- Currat M., Excoffier L. Modern humans did not admix with Neanderthals during their range expansion into Europe. PLoS Biol. 2004;2:e421.
- Leakey L. S. Homo habilis, Homo erectus and the Australopithecines. Nature. 1966;209:1279-1281.
- Oeppen J., Vaupel J. W. Demography. Broken limits to life expectancy. Science. 2002; 296:1029-1031.
- Pilbeam D. The descent of hominoids and hominids. Sci. Am. 1984;250:84-96.
- Pilbeam D. Hominoid systematics: the soft evidence. Proc. Natl. Acad. Sci. U S A. 2000; 26;97:10684-10686.
- Robinson J. T. The distinctiveness of Homo habilis. Nature. 1966;209:957-960.
- Strait D. S., Grine F. E. Inferring hominoid and early hominid phylogeny using craniodental characters: the role of fossil taxa. J. Hum. Evol. 2004;47:399-452.
- Vaupel J. W., Carey J. R., Christensen K., Johnson T. E., Yashin A. I., Holm N. V., Iachine I. A., Kannisto V., Khazaeli A. A., Liedo P., Longo V. D., Zeng Y., Manton K. G., Curtsinger J. W. Biodemographic trajectories of longevity. Science. 1998;280:855-860.
- Watson E., Forster P., Richards M., Bandelt H. J. Mitochondrial footprints of human expansions in Africa. Am. J. Hum. Genet. 1997;61:691-704.
- Wolpoff M. H. Lucy’s lower limbs: long enough for Lucy to be fully bipedal? Nature. 1983;304:59-61.
- Wood B. Human evolution. Bioessays. 1996;18:945-954.