Tema 7. Circulación capilar
La circulación capilar desarrolla la función básica y última del sistema cardiovascular: el intercambio de sustancias entre la sangre y las células del organismo facilitando su supervivencia.
7.1 Estructura y tipos de casos capilares
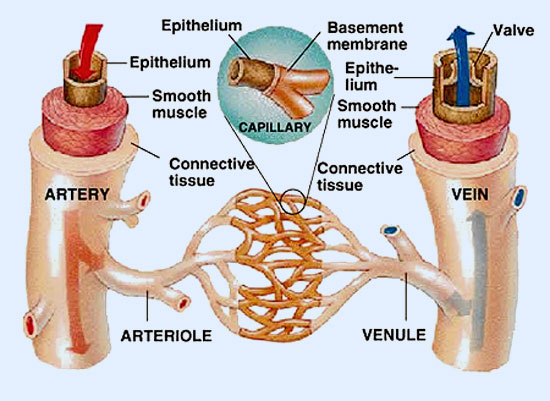
Los capilares son las ramas más pequeñas del árbol circulatorio, y son el centro del sistema de la microcirculación. De la porción final de las arteriolas, se originan las metaarteriolas, punto de origen de los capilares. Éstas, con un diámetro de 10-20 μ, conservan fibras de músculo liso situadas de forma discontinua. En su punto de origen, los capilares están rodeados de un anillo de músculo liso denominado esfínter precapilar, éste se contrae o relaja determinando el flujo existente en un lecho capilar concreto, aunque no están presentes en todos los territorios.
Por término medio, un capilar mide aproximadamente 1 mm de longitud y su diámetro está entre 5 y 10 m, dejando pasar justo un eritrocito (que en muchos casos ha de hacerlo deformándose). Se estima que en un adulto existen unos 40.000 millones de capilares, lo que en función de longitud individual, daría unos 40.000 km de capilares, esto supone una superficie total disponible para el intercambio de 700 m2. En su conjunto, representa el mayor sistema de comunicación entre las células y la sangre. Ninguna célula viva se encuentra alejada más allá de 20 micras de un capilar, distancia suficientemente pequeña para que el intercambio resulte extraordinariamente eficiente.
El flujo sanguíneo en el lecho capilar es el 5% del gasto cardíaco y depende estrechamente del resto de los vasos que integran la microcirculación. La estructura de los capilares varía de órgano a órgano, pero típicamente están formados por una capa única de células endoteliales apoyadas sobre una membrana basal. Carecen de músculo liso y de fibras elásticas.
7.1.1 Estructura de la pared capilar
Todo el sistema circulatorio está interiormente tapizado por una capa de células de epitelio escamoso que, en conjunto, reciben el nombre de endotelio. La pared capilar está formada por una sola capa de células endoteliales y una membrana basal externa.
Las células endoteliales presentan espacios intercelulares entre ellas por los que pueden pasar sustancias susceptibles de intercambio. El número y tamaño de estos canales, poros o fenestraciones es muy variable y depende del tipo de capilares analizado. Según la forma y la cantidad de estos poros, los capilares se clasifican en:
- a) Capilares continuos. Son los más abundantes, presente en muchos lechos vasculares. Presenta uniones estrechas abundantes. Sus células firmemente adosadas entre sí, apenas dejan espacios entre ellas, y hacen que la zona sea prácticamente impermeable.
- b) Capilares fenestrados. Como indica su nombre, la presencia de ventanas o fenestraciones es su rasgo más característico. Estos poros pueden estar obturados o no por un diafragma. Estos rasgos permiten decir que a través de estos capilares pasan sustancias de mayor peso molecular que las que lo hacen a través de los capilares continuos.
- c) Capilares discontinuos o sinusoides. En este tipo de capilares, la pared está completamente interrumpida, dejando grandes espacios entre las células endoteliales.
7.2 Intercambio a nivel capilar
Cuando la sangre alcanza el lecho capilar, la velocidad con que circula por el interior de estos vasos es muy baja. Ello es debido a que en un sistema de tubos que se ramifica, la velocidad disminuye proporcionalmente al aumento de la sección conjunta de las ramificaciones. Así en la aorta, de sección pequeña, la velocidad es muy grande (400 mm/seg), va descendiendo a nivel de arterias y arteriolas y a nivel capilar se hace mínima (0,1 mm/seg). Este dato junto con la delgadez de la pared capilar proporciona las condiciones de tiempo y espacio necesarias para que el intercambio pueda efectuarse de la manera más óptima posible.
7.2.1 Tipos de intercambio entre capilares y líquido intersticial
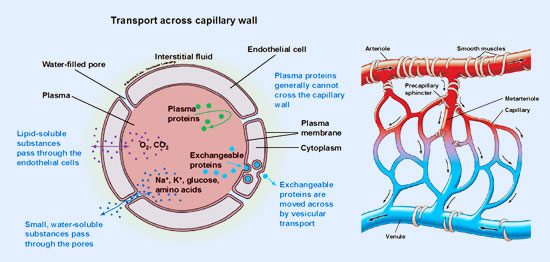
Existen tres modalidades de transporte a través de la pared capilar: difusión, pinocitosis o transporte vesicular y ultrafiltración o reparto de líquidos.
- a) Difusión. Es el mecanismo más importante de los tres. Es un tipo de transporte pasivo, regulado por la ley de Fick (Cap. Fisiología Celular). Todos los intercambios entre la sangre y las células utilizan como vía intermediaria el líquido intersticial. Los gradientes para la correcta difusión de gases respiratorios, nutrientes y productos de desecho se crean por el metabolismo celular que consume unos y produce otros, dando lugar a aumentos o disminuciones de su concentración en el líquido intersticial. El metabolismo es, por tanto, la causa de la creación de gradientes y del movimiento de las moléculas a uno y otro lado de la pared capilar.
- b) Pinocitosis o transporte vesicular. Las vesículas permiten realizar procesos de endo y exocitosis constituyendo verdaderos canales transcelulares de transporte.
- c) Ultrafiltración. Reparto de líquidos. Este tipo de transporte tiene como función básica la redistribución de líquidos extracelulares. El líquido extracelular está formado básicamente por dos componentes: el plasma, con un volumen de 3 litros, y el líquido intersticial, con un volumen aproximado de unos 10 litros. El líquido intersticial puede ser utilizado como reservorio o almacén, pudiendo recibir líquido del plasma o bien proporcionándoselo al mismo.
El plasma tiene dos tipos de solutos:
- Solutos de bajo peso molecular, que atraviesan sin ninguna dificultad la pared capilar y, por tanto, tienen la misma concentración a ambos lados.
- Solutos de alto peso molecular o coloides: las proteínas, incapaces de atravesar la pared y que se encuentran en elevada concentración dentro del capilar (6-8 g/100ml), y baja en el líquido intersticial (0,7-2 g/100ml).
La magnitud de este flujo y el hecho de que el capilar no se vacíe viene determinado por las fuerzas que intervienen en el equilibrio de Starling. A través de este equilibrio, existe un movimiento de líquido sin modificación de las concentraciones de solutos de bajo peso molecular, a un lado y otro. Sin embargo, las cantidades (volumen) tanto de agua como de solutos cambiarán en función de la dirección preferente del flujo.
7.2.2 Equilibrio de Starling
Este equilibrio describe como se mantienen los volúmenes distribuidos correctamente. Los movimientos de agua en el lecho capilar se desarrollan en ambos sentidos, mediante dos sistemas de fuerzas opuestas, las presiones hidrostáticas y coloidosmóticas, que se establecen a través de la pared capilar.
Existen dos gradientes contrarios de presión que son: uno hidrostático, ejercido por el líquido, y otro coloidosmótico dependiente de las proteínas. De acuerdo con estas diferencias, el movimiento depende de cuatro variables individuales que son las siguientes:
- Presión hidrostática capilar (Phc). Cuando la sangre llega al extremo arterial del capilar la presión hidrostática o arterial es de 35 mm de Hg. y, como el capilar también ofrece cierta resistencia al flujo, la presión sigue descendiendo y en el extremo venoso del mismo la presión ha caído a 16 mm Hg.
- Presión hidrostática intersticial (Phi). Es la presión que ejerce el líquido intersticial. Su medida es compleja y se asume que su valor es 0 mm Hg., aunque en algunos tejidos se han obtenido valores subatmosféricos o negativos que oscilaban entre –3 y –9 mm Hg., debido probablemente al drenaje linfático.
- Presión osmótica, coloidosmótica, u oncótica capilar (pC). Es la presión desarrollada por las proteínas plasmáticas. El efecto osmótico de estos solutos empuja al agua hacia el interior del vaso. Su valor es de aproximadamente 28 mm Hg.
- Presión osmótica o coloidosmótica intersticial (pI). Es la presión que ejercen las proteínas del líquido intersticial. Como su concentración es mucho más baja que la plasmática (1-2 g/dL), su valor es de 3 mm Hg.
La combinación de estas cuatro presiones a lo largo del recorrido capilar permite analizar los movimientos, totalmente pasivos de líquidos, que tienen lugar y que son dependientes de la siguiente ecuación:
PF = (Phc + pI) – (Phi + pC)
Donde Pf se define como presión eficaz o neta de filtración y se define como la diferencia de presiones que empujan el líquido hacia fuera (Phc + pI), y las presiones que empujan hacia adentro (Phi + pC).
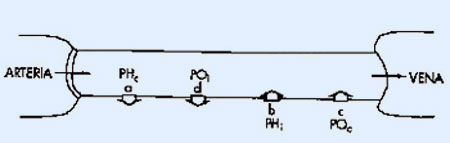
- a) Flujo en el extremo arterial del capilar. En esta región el sumatorio de presiones que empujan el líquido hacia el exterior del capilar es superior al sumatorio de las que lo empujan hacia el interior. La diferencia da como resultado una presión eficaz de filtración de 10 mm Hg. Por lo tanto en esta primera porción del capilar se produce la filtración o salida de líquido hacia el espacio intersticial.
- b) Flujo en el extremo venoso del capilar. En este punto la diferencia de presiones se invierte siendo el sumatorio de presiones hacia el exterior inferior al sumatorio de presiones hacia el interior. El resultado final da una presión eficaz de filtración de –9 mm Hg, lo que indica una "filtración inversa" o movimiento de líquido hacia el interior del vaso definido como absorción o reabsorción.
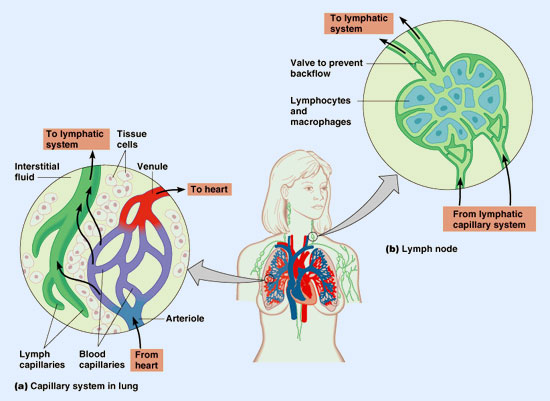
Así, ambos procesos tienden a compensarse; sin embargo, debido a que las presiones netas no son iguales existe un flujo mayor de filtración que de absorción. En términos porcentuales, del 100% de filtrado (20 litros/día), se absorbe el 90% (18 litros/día) a través de la región venosa del capilar, y el 10% restante (2 litros/día) será recogido por el sistema linfático.
Una cuestión que debe remarcarse es que la filtración y absorción a nivel capilar no desempeñan un papel significativo en el intercambio de nutrientes y productos de desecho, ya que la principal fuerza de empuje para ambos, no es el gradiente de presiones, sino el gradiente de concentraciones. Comparativamente las cantidades que se mueven, a través de filtración y absorción, son muy pequeñas.