8.4 Efectos de los desastres de origen humano
Las transformaciones más importantes inducidas en la Biosfera por el ser humano han sido causadas por sus asentamientos, infraestructuras y actividades económicas (agricultura, ganadería, explotación de recursos…). Sus primeras manifestaciones son muy antiguas ya que remontan a la prehistoria aunque desde aquel momento no han cesado de intensificarse y de extenderse de manera que hoy afectan a la práctica totalidad de la tierra. Evidentemente los grupos humanos no han perseguido la destrucción gratuita de la naturaleza sino que los impactos, de los que no siempre se ha tenido conciencia, han sido la consecuencia inevitable (o considerada como tal) de sus actividades.
Esta situación ha producido cambios sin precedentes en la historia de la Biosfera y ha supuesto la extinción de numerosas especies. Sin embargo, al ser fruto de una evolución muy prolongada en el tiempo, tanto los distintos ecosistemas como muchas especies se han podido ir adaptando a la presión humana originando los ambientes seminaturales que hoy dominan la mayor parte de la superficie terrestre.
 |
|
El ser humano crea con creciente frecuencia situaciones que causan la muerte de la mayor parte de las especies o que dan lugar a ambientes hostiles y aparentemente incompatibles con la vida. Sin embargo, ésta acaba siempre por volver a estos lugares aunque los ecosistemas resultantes no sean siempre idénticos a los preexistentes. Foto: rebecos en las minas de los Puertos de Áliva (Cantabria, España). |
Pero, a medida que el nivel tecnológico de nuestra especie ha ido aumentando, también lo ha hecho la posibilidad de provocar terribles desastres capaces de destruir cualquier forma de vida o alterar gravemente la distribución de los organismos en muchos kilómetros a la redonda. Estos acontecimientos son imprevisibles, prácticamente instantáneos, pueden afectar a cualquier punto de la Biosfera y, dada su inmediatez, no permiten una adaptación previa de los seres vivos por lo que sus consecuencias en el medio natural pueden prolongarse durante largos periodos de tiempo.
Por supuesto, los acontecimientos catastróficos son consustanciales a la naturaleza y han contribuido a la evolución de las especies y a la diferenciación de muchos ecosistemas. Los incendios forestales, erupciones, inundaciones catastróficas o, incluso, el impacto de meteoritos, por no citar más que algunos ejemplos, han estimulado la evolución de la Biosfera y forman parte de nuestra historia. Sin embargo, la interacción entre los sistemas humanos y naturales está incrementando exponencialmente el número de desastres “naturales” e industriales y generando nuevas situaciones de riesgo que, por su recurrencia creciente, podrían convertirse en serios condicionantes de la distribución de las especies en un futuro próximo.
Sabemos que la vida posee cualidades extraordinarias que le permiten superar con éxito estas crisis e incluso que cada una de ellas facilita su perfeccionamiento. Sin embargo, es frecuente que el “antes” y el “después” no sean iguales y que la recuperación tras el desastre sea complicada e introduzca factores de distorsión en los ecosistemas, algo que, dada la precaria situación de muchos de ellos, es imprescindible conocer. Para ello se va a analizar a continuación la respuesta de la Biosfera a tres tipos de situaciones desastrosas producidas por la actividad humana. Los ejemplos posibles son muchos más pero creemos que los casos presentados son suficientemente ilustrativos y permiten cumplir el objetivo perseguido.
Los incendios forestales
Los incendios forestales son fenómenos bastante comunes en la naturaleza aunque su distribución es muy desigual dependiendo básicamente del clima. Pueden ser debidos a múltiples causas tales como la caída de un rayo (que es la más frecuente), erupciones volcánicas, procesos de combustión espontánea en presencia de gases inflamables o, incluso, a una chispa producida por la caída de algún objeto.
 |
|
Los incendios causan graves alteraciones en los ecosistemas ya que destruyen la cubierta vegetal, dañan los suelos y matan a los animales con menor capacidad de desplazamiento. Las aves y grandes vertebrados suelen librarse de las llamas aunque pierden sus hábitats y fuentes de recursos habituales y se verán obligados a desplazarse a otras zonas durante los meses o años posteriores a cada siniestro. Foto: ciervos protegiéndose del calor de un incendio en Bitterroot National Forest (Montana- EEUU). Imagen de dominio público disponible en http://upload.wikimedia.org/wikipedia/commons/d/d8/Deerfire_high_res_edit.jpg. |
{kind=link}
El fuego ya existía sobre la superficie terrestre cuando aparecieron los primeros bosques y a lo largo del tiempo tanto las especies, de forma individual, como los ecosistemas de los que forman parte se han ido adaptando a él de distintas formas. El fuego ha sido un factor esencial en la evolución de manera que las regiones más propicias a los incendios por razones climáticas suelen estar ocupadas por formaciones naturales que soportan bien el paso esporádico de las llamas y que, incluso, lo necesitan para mantenerse en condiciones óptimas. De ahí que, siempre que su frecuencia e intensidad no se alejen excesivamente de las propias del medio natural, los incendios, lejos de constituir un problema, sean un factor más de los ecosistemas y contribuyan eficazmente a su diversidad.
Sin embargo, la situación actual es muy distinta ya que se estima que cada año el fuego destruye unos 350 millones de ha de vegetación en todo el mundo y que cerca del 90% de todos esos incendios es provocado. Es cierto que la vegetación siempre consigue regenerarse, unas veces deprisa, otras con más dificultad, pero en un número creciente de lugares la repetición es excesivamente rápida y la cubierta vegetal no logra alcanzar nunca sus estadios maduros lo que, a la larga, conlleva cambios que terminan siendo irreversibles.
 |
|
Tras los incendios más intensos, la vegetación queda destruida y desaparece todo signo de vida de la superficie. Sin embargo, bastan algunas horas para que acudan atraídos por el calor los primeros insectos o un tipo peculiar de geógrafos capaces de deambular por estos medios manteniendo su ropa impoluta. Foto: superficie de matorral destruida por un incendio en Las Rozas (Cantabria, España). |
Los vegetales carecen de sistemas activos de defensa contra el fuego y permanecen fijos en el suelo por lo que, cuando se produce un incendio, no pueden librarse de sufrir sus efectos. Sin embargo, la adaptación de las plantas a esta eventualidad es muy desigual lo que, unido a su diferente capacidad de respuesta, permite clasificarlas en varios grupos:
- Especies intolerantes al fuego: generalmente son muy combustibles y quedan totalmente calcinadas por el incendio.
- Especies germinadoras: aunque no soportan el fuego garantizan su supervivencia produciendo abundantes semillas cuya dispersión y germinación son favorecidas por el propio incendio.
- Especies rebrotadoras: toleran relativamente bien el fuego y tras el incendio se recuperan emitiendo nuevos brotes desde sus raíces o ramas. Suelen ser geófitas, que crecen muy deprisa gracias a las reservas contenidas en sus raíces, o árboles en los que el fuego solo daña las partes más externas sin perjudicar gravemente a los órganos vitales.
- Especies resistentes al fuego: son aquellas que no suelen sufrir daños importantes y que gracias a ello no ven comprometida su supervivencia. La mayoría son grandes árboles cuyas copas quedan fuera del alcance del fuego y que se encuentran protegidos por gruesas cortezas aislantes.
Como las especies resistentes al fuego son muy escasas, la verdadera competencia tras un incendio se produce entre las rebrotadoras y las germinadoras. Sin embargo, las primeras tienen ventaja ya que les basta regenerar sus partes aéreas para poder ocupar el terreno después del fuego mientras que las germinadoras necesitan mayor tiempo para crecer y alcanzar la madurez.
 |
|
Las áreas sin quemar y los árboles supervivientes al fuego facilitan mucho la regeneración posterior de la cubierta vegetal ya que garantizan un aporte de semillas y se convierten en un foco de atracción de animales. Foto: Área recorrida por un incendio en Las Rozas (Cantabria, España). |
La recuperación de la cubierta vegetal depende, por un lado, de la tolerancia al fuego de las plantas de cada lugar (favoreciendo la expansión de las llamadas “pirófitas” o mejor adaptadas) y, por otra, de las condiciones ambientales posteriores al incendio tales como la calidad del suelo, iluminación, disponibilidad de agua, temperatura y características de las áreas circundantes no quemadas. En general los ecosistemas maduros y mejor conservados arden más difícilmente y su recuperación posterior suele ser más rápida que los entornos más degradados o afectados por fuegos repetidos.
Inmediatamente después del incendio la superficie afectada no muestra signos aparentes de vida. Sin embargo la combustión implica la mineralización de la materia orgánica y, por tanto, un fuerte aporte de nutrientes al suelo. Al mismo tiempo, el fuego estimula la germinación de las semillas de numerosas especies y crea condiciones que en un primer momento facilitan el desarrollo de las plantas. Gracias a ello, en el plazo de unos días o semanas éstas empiezan a reaparecer. Las primeras en hacerlo son leñosas rebrotadoras, helechos y algunas herbáceas perennes que se benefician del momentáneo aumento de la fertilidad. En algunos casos esta etapa es aprovechada también por especies oportunistas o invasoras para instalarse en territorios en los que previamente no se encontraban gracias a su gran capacidad de colonización y a la ausencia de competidores.
Posteriormente surgirán las herbáceas anuales, que deben esperar el momento adecuado para germinar, y el resto de las especies preexistentes, primero las germinadoras y, por fin, cuando hay un aporte suficiente de semillas desde las áreas vecinas, las más intolerantes. No obstante, la recuperación total del ecosistema y la vuelta a una situación comparable a la inicial requiere bastante tiempo y en las zonas más humanizadas es frecuente que quede interrumpida por la ocurrencia de nuevos incendios que obligan a reiniciar el ciclo sin permitir que éste alcance sus últimos estadios.
Las especies implicadas y la velocidad de recuperación son diferentes en cada ambiente y dependen de la severidad del incendio y de la supervivencia de la biomasa aérea pero la secuencia que se observa es siempre del mismo estilo.
En la Cordillera Cantábrica, por ejemplo, tras la quema de un brezal-escobal en un área de pastos bastan algunos días para que hagan su aparición los helechos (Pteridium aquilinum), favorecidos por las reservas contenidas en sus grandes rizomas, las primeras hojas de las plantas más resistentes como las zarzas (Rubus spp) y algunas gramíneas. Durante los primeros años posteriores al incendio se produce un aumento de la diversidad florística entre las herbáceas (gramíneas, liliáceas…), las más beneficiadas por los efectos favorables del fuego, y sólo la progresiva recuperación del matorral, dominado por las rebrotadoras primero (Erica australis, Arctostaphylos uva-ursi, Halimium alyssoides…) y por las germinadoras después (Calluna vulgaris, Erica umbellata) pone fin a esta momentánea riqueza del estrato herbáceo y restablece una situación similar a la preexistente..
 |
|
Algunas plantas tienen la capacidad de rebrotar rápidamente tras un fuego y de beneficiarse del aporte de fertilizantes proporcionados por las cenizas. Foto: rebrote de una zarza (Rubus sp) a las tres semanas de un incendio en Cantabria. |
El fuego genera una situación de estrés muy importante por lo que, en caso de repetirse, acaba por convertirse en un significativo factor de evolución tanto para los ecosistemas como para las especies existentes en ellos. Eso permite que en los ambientes en los que los incendios naturales resultan habituales se puedan observar numerosos mecanismos de adaptación en las plantas e incluso una particular fisonomía de las masas vegetales dominadas por las especies pirófitas.
En latitudes medias abundan las plantas capaces de soportar fuegos reiterados de baja intensidad gracias una gruesa corteza que actúa de aislante térmico. Es el caso del alcornoque (Quercus suber), del abeto de Douglas (Pseudotsuga Menziesii), o del pino canario (Pinus canariensis).
 |
|
Las plantas pirófitas desarrollan diversas estrategias para soportar el fuego y aprovecharlo en su competencia con las demás especies. En la foto, tomada en Fuencaliente (La Palma-Canarias, España), se observa como millares de brotes de pino canario (Pinus canariensis) recubren una superficie recién quemada. |
En otros casos, la estrategia está relacionada con la dispersión de las semillas. A este grupo pertenecen numerosas coníferas que tienen conos serotinos (piñas de apertura retardada). Al iniciarse el incendio, las semillas permanecen protegidas dentro de los conos y adheridas a ellos por la resina pero, durante el transcurso mismo, el calor funde la resina, las escamas se separan abriendo la piña y las semillas quedan liberadas. De este modo podrán dispersarse durante los días siguientes, cayendo sobre una capa de cenizas idónea para su germinación y sin ningún rival que pueda limitar su crecimiento.
Por otra parte, el calor estimula la germinación de las semillas de muchas especies que, como Genista florida, Cytisus scoparius, Cistus ladanifer o Cistus laurifolius son muy comunes en muchos montes gracias, precisamente, a su capacidad para germinar rápidamente tras el paso del fuego.
Por fin, hay un último grupo que se beneficia indirectamente del fuego: se trata de las heliófilas, necesitadas de una iluminación intensa para poder desarrollarse y que, en condiciones normales, tienen dificultades para prosperar dada la penumbra reinante dentro del bosque.
La repetición de los incendios tiene importantes consecuencias para la cubierta vegetal ya que explica la presencia o ausencia de numerosas especies y da lugar a modelos de sucesión que determinan un complejo mosaico de edades y de comunidades vegetales. En las regiones con incendios frecuentes, la vegetación intolerante al fuego termina desapareciendo, desplazada por la mejor adaptada (que, muchas veces, es además muy inflamable). Por eso, una de las consecuencias más insidiosas de los incendios es que crean condiciones que facilitan la reaparición de fuego en los años siguientes alimentando un círculo vicioso cada vez más cerrado.
A la larga, si la repetición de los incendios sigue a un ritmo tal que la recuperación de los árboles no resulta posible, las formaciones boscosas son sustituidas por otras más inmaduras, preforestales, arbustivas o incluso herbáceas. Es el caso de gran parte de los brezales o escobales del Norte de España, de los jarales mediterráneos o de las grandes extensiones de gramíneas de las sabanas tropicales cuya existencia se debe en gran medida a los incendios. En caso extremo, se puede llegar a la aparición de estepas o semidesiertos aunque, a la inversa, si quedan retazos forestales que permitan un aporte suficiente de semillas, la vegetación de estos lugares tiende a progresar evolucionando espontáneamente hacia estadios más complejos en cuanto los incendios desaparecen o logran ser controlados.
 |
|
El uso recurrente del fuego puede acarrear una transformación irreversible de la vegetación que suponga la sustitución de los bosques por formaciones abiertas de matorral, sabana o herbáceas. Amplias zonas de sabana o estepa que hoy consideramos naturales en todo el mundo son, en realidad, formaciones de sustitución generadas por los incendios. Foto: Kata Tjuta (Territorios del Norte, Australia). |
A diferencia de la vegetación, los animales perciben el fuego y son capaces de reaccionar antes de la llegada de las llamas. Sin embargo, sus respuestas son muy dispares y no siempre acertadas: muchos vertebrados huyen del peligro, otros animales intentan refugiarse en madrigueras o escondites mientras que un tercer grupo, entre los que se encuentran numerosos insectos, resultan atraídos por el fuego y se precipitan literalmente dentro de él. Es evidente que la eficacia de estas respuestas depende de la capacidad de las distintas especies para ponerse a salvo, y si bien la mayoría de los grandes mamíferos o de las aves adultas pueden librarse de las llamas, los animales de menor movilidad que no pueden separarse del suelo, como los reptiles, anfibios o la mayor parte de la microfauna terrestre, resultan mucho más vulnerables y soportan mal los incendios repetidos.
No obstante, aunque muchos de ellos puedan escapar y salvar la vida, el fuego perjudica durante bastante tiempo a los animales ya que destruye la vegetación que les es necesaria para vivir. Por eso, suelen tardar bastante en retornar definitivamente al área afectada.
Las consecuencias sobre la microfauna son insuficientemente conocidas aunque el impacto es particularmente grave en la del suelo ya que dificulta la vuelta a la normalidad de los procesos edafogenéticos y reduce los recursos alimenticios de los que dispondrán numerosos omnívoros y carnívoros.
En general el número de insectos disminuye drásticamente tras la ocurrencia de un fuego aunque éste también favorece a algunas especies que proliferan durante los años posteriores al incendio gracias a la acumulación de madera muerta y a la momentánea desaparición de sus depredadores.
En todos los casos, la colonización de las zonas quemadas por la fauna es más fácil cuando subsisten áreas próximas inalteradas o cuando la vegetación forma un mosaico de piezas muy recortadas como consecuencia de la repetición de sucesivos fuegos. En tales ocasiones, coexisten manchas de vegetación con distintos grados de madurez o alteración y cada especie dispone de hábitats favorables en los que refugiarse y desde los que iniciar una posible expansión posterior. En cambio, los grandes incendios que arrasan miles de hectáreas impiden el mantenimiento de estas áreas refugio y crean condiciones uniformes en grandes extensiones, lo que dificulta la recolonización “de fuera a dentro” y hace necesarios periodos de tiempo más prolongados hasta la total desaparición de las consecuencias del siniestro.
Por fin, la velocidad de la recolonización depende mucho también de la capacidad de desplazamiento de los distintos tipos de animales presentes en cada región y aunque los de mayor talla, pertenecientes habitualmente a los niveles superiores de las cadenas tróficas, pueden reaparecer muy deprisa, es normal que los pertenecientes a niveles inferiores encuentren mayor dificultad para hacerlo.
 |
|
Algunos árboles disponen de toda una panoplia de recursos para soportar incendios repetidos. Es el caso del pino canario (P.canariensis) que se beneficia de su gran altura y corteza aislante, es capaz de rebrotar tras el siniestro y cuyas semillas se dispersan y germinan de manera óptima en un terreno recién quemado. Foto: aspecto de un pinar dos años después de un incendio en el monte la Esperanza (Tenerife-Canarias, España). |
Los efectos del fuego en la fauna se producen a veces al nivel de las poblaciones, habiéndose observado anomalías en cadena en la abundancia y en el éxito reproductor de determinadas especies tras un incendio. La población de aves, por ejemplo, acusa muy rápidamente los cambios en la abundancia de recursos tróficos, refugio y condiciones para la nidificación (y ello a pesar de presentar cierta plasticidad en relación con el uso del hábitat y con la localización del nido). Por eso, numerosas rapaces se ven favorecidas por los incendios: las presas son más fáciles de localizar y su número aumenta gracias al incremento de la vegetación herbácea o de los insectos que les sirven de alimento. Pero esta situación es pasajera ya que pronto harán su aparición las plantas leñosas que, acompañadas por otros insectos, constituirán recursos distintos que serán aprovechados por otros animales y así sucesivamente.
Es evidente que en los lugares en los que los incendios se repiten con suficiente frecuencia e intensidad como para alterar de forma irreversible la cubierta vegetal, la transformación de los hábitats puede acarrear modificaciones más o menos definitivas de las comunidades de fauna. La destrucción por el fuego de los bosques tropicales y su sustitución por sabanas, por ejemplo, supone la desaparición de los roedores arborícolas y frugívoros, de ciertos antílopes, simios y, seguramente, de insectos asociados a las formaciones iniciales.
Consecuencias de las mareas negras
Cada año se producen en el mundo un buen número de accidentes que implican vertidos masivos de hidrocarburos en el mar. Unas veces son causados por el hundimiento de buques, en ocasiones superpetroleros que cargan varios cientos de miles de toneladas de crudo, otras veces por accidentes o fugas en plataformas de extracción, otras más por vertidos criminales o prácticas inadecuadas. Además, existe una contaminación “de fondo” originada por escapes naturales desde el sustrato geológico. La sustancia que se derrama es distinta en cada caso ya que existen muchos tipos diferentes de petróleo y la variedad de sus productos derivados es aún mayor. Sin embargo, el resultado de estos derrames, que se extienden rápidamente por amplias superficies tras formar una fina película flotante, se parece siempre e incluye una grave contaminación de la línea de costa y de la superficie marina y la muerte de la mayoría de los animales, plantas y algas alcanzados por ellos.
 |
|
Los vertidos de petróleo en el mar forman una lámina flotante que se desplaza impulsada por las olas y corrientes contaminando todo lo que encuentra a su paso. Muy tóxica y difícil de limpiar, produce la muerte de la mayoría de los animales, plantas o algas que quedan recubiertos por ella. Foto: pato recubierto de petróleo durante una marea negra. Imagen de dominio público disponible en: http://upload.wikimedia.org/wikipedia/commons/3/3f/Oiled_bird_3.jpg. |
{kind=link}
De forma complementaria, los hidrocarburos vertidos sufren diversas reacciones de oxidación que suponen el agotamiento del oxígeno disuelto en las capas más superficiales de agua y producen la muerte por asfixia de numerosos organismos.
Pero los efectos ambientales de las mareas negras no se limitan al episodio de muertes directamente asociadas al contacto con el petróleo sino que se extienden al conjunto del ecosistema y persisten a lo largo de cierto tiempo de varias maneras:
- Degradación de los biotopos y de los ecosistemas causada por la asfixia del medio y la destrucción directa de los hábitats de numerosos organismos.
- Alteración de las biocenosis por contaminación de las redes tróficas y bioacumulación en los organismos situados en los niveles superiores.
- Daños a largo plazo como consecuencia de enfermedades o deficiencias inducidas en el metabolismo o en los sistemas inmunitario, reproductor u otros de muchos animales.
Aunque no son las únicas afectadas por el desastre, los daños más conspicuos y que más impresionan a la opinión pública, son los sufridos por las aves. Éstas, al quedar recubiertas de petróleo, pierden capacidad de desplazamiento, flotan más difícilmente y quedan peor aisladas del frío. Además, los hidrocarburos se acumulan en su sistema respiratorio produciéndoles asfixia por lo que terminan muriendo al cabo de algunos días.
Cuando el vertido se produce en alta mar sus consecuencias son menos graves que cuando alcanza el litoral ya que en éste último caso la mayor parte de las especies viven ancladas a la superficie rocosa y no tienen la capacidad de escapar de las de mar abierto.
Sin embargo, pasada la primera fase del desastre, la cantidad de petróleo flotante va disminuyendo gracias a diversos procesos naturales (evaporación, fotooxidación y posterior disolución, sedimentación en el fondo, dispersión, biodegradación…) de eficacia muy desigual dependiendo del tipo de petróleo y del clima exterior pero también gracias a las posibles intervenciones humanas de recogida y limpieza. A partir de este momento se hace posible un progresivo retorno de la vida.
 |
|
Las mareas negras causan una grave contaminación que acaba con cualquier forma de vida existente en la franja de costa alcanzada por ellas. Foto: playa recubierta de petróleo por la marea negra del Exxon Valdez en Disk lsland (Prince William Sound- Alaska, EEUU). Imagen de dominio público disponible en: http://www.flickr.com/photos/arlis-reference/4750349750/sizes/o/in/photostream/. |
Normalmente, hacen falta unos dos años para que el litoral recupere cualitativa y cuantitativamente un estado más o menos comparable al preexistente lo que, en comparación con otro tipo de desastres, es un plazo muy breve. No obstante, en lugares desfavorables por su inaccesibilidad, clima y escasa agitación o renovación del agua pueden llegar a ser necesarias varias décadas para que la recuperación alcance un nivel aceptable.
De hecho, la eficacia de la recolonización biológica del litoral varía mucho dependiendo tanto del tipo y cantidad de petróleo extendido como de las características del litoral afectado.
En general, cuanto mayor es la fracción ligera del vertido, más capacidad tiene el producto para infiltrarse en el organismo y producir un envenenamiento (aunque, en contrapartida, estos productos ligeros son también los que se evaporan o biodegradan más fácilmente). A la inversa, los fueles muy densos son más persistentes aunque se extienden peor (dejando zonas limpias que facilitarán la recolonización posterior), y resultan menos tóxicos.
Por otra parte, el petróleo depositado en un tramo de costa expuesto a fuertes mareas y oleaje va a estar sometido a continuos cambios de temperatura, de humedad, etc. que favorecen su descomposición contrariamente a lo que ocurre en litorales con menos energía.
La recolonización suele verificarse por etapas, empezando por los taxones más simples y resistentes y terminando por los animales más evolucionados pero de hábitos preferentemente sedentarios, como algunos grandes crustáceos. En todos los casos, estos organismos proceden de las áreas vecinas no contaminadas y su desplazamiento está favorecido por la continua agitación del medio marino.
Los primeros en volver después del desastre son pequeños organismos oportunistas de ciclo corto que se benefician del vacío creado por la muerte de sus competidores habituales y que son capaces de soportar elevados niveles de toxicidad (plancton, algas verdes, poliquetos…). Algunas de estas especies logran incluso beneficiarse del petróleo aprovechando el exceso de carbono y de hidrógeno que libera su degradación. Es el caso de las algas verdes que, a veces, experimentan una proliferación explosiva durante un corto plazo y que de este modo proporcionan abundante alimento a los organismos marinos que se alimentan de ellas facilitando su retorno (que, a su vez, permitirá el de las especies de los niveles tróficos superiores).
De hecho, no es raro que tras una marea negra se produzca una auténtica y aparentemente contradictoria explosión de vida. Ello es debido a una distorsión transitoria de las cadenas alimentarias pero el hecho desaparece rápidamente cuando se recupera el equilibrio preexistente.
Sin embargo también pueden producirse complicaciones insidiosas capaces de perturbar durante varios años el proceso de recolonización: ciertos hidrocarburos originan problemas reproductivos o tumores en las especies más expuestas a ellos como los moluscos filtradores (mejillones, ostras…) o los animales situados en los niveles superiores de las cadenas tróficas (mamíferos marinos…). Su acumulación entre los cantos de las playas o fondos marinos puede prolongar anormalmente una situación de toxicidad y generar en muchos animales problemas crónicos de salud causantes de malformaciones, crecimiento anómalo o diversas lesiones que, a veces, han sido observados más de una década después del desastre.
Todo lo anterior nos demuestra la gran capacidad de recuperación de los medios costeros y el carácter transitorio de la mortandad causada por las mareas negras pero, también, el riesgo que supone la repetición continua de estos episodios en algunos tramos particularmente expuestos del litoral en los que la recuperación total se hace cada vez más difícil y más lenta y la degradación ambiental termina siendo semipermanente.
 |
|
En mares agitados y no excesivamente fríos la descomposición del petróleo y recuperación de la vida son bastante rápidos y suelen producirse en el plazo de algunos años. Foto: restos muy degradados de petróleo de la marea negra del Prestige en la costa de Cantabria (España). |
Efectos de una catástrofe nuclear: Chernobil
Un tipo de catástrofe totalmente distinta a las anteriores es la causada por la liberación repentina a la atmósfera de grandes cantidades de radiactividad. Hasta el momento este hecho sólo se ha producido como consecuencia de la explosión de bombas nucleares (Hiroshima y Nagasaki pero, también Mururoa, Bikini, Kazakhstan, Nevada u otros “polígonos de ensayos al aire libre”) y a causa de grandes accidentes en instalaciones civiles (Fukushima, Chernobil, Kyshtym, Windscale, Three Miles Island, etc).
Dadas las extremas medidas de seguridad que rodean las instalaciones nucleares y sus materiales estos acontecimientos son mucho más improbables que los descritos hasta aquí. Sin embargo, la extrema gravedad que pueden revestir para la humanidad, la relación existente entre la tecnología nuclear y las armas y el sistemático intento de ocultación que suele producirse tras cualquier incidente, han hecho que para gran parte de la opinión pública los accidentes nucleares (que, por otra parte, son muy difíciles de comprender para el profano y siempre tienen algo de “misterioso”) se hayan convertido en el paradigma del desastre inducido por el ser humano y un motivo permanente de preocupación.
Sin embargo, y contrariamente a lo que se suele pensar, el impacto de la radiactividad en el medio natural está menos claro y plantea no pocas dudas y contradicciones.
El caso mejor documentado, que utilizaremos como ejemplo, es el accidente que se produjo el 20 de abril de 1986 cuando uno de los reactores de la central nuclear de Chernobil (Ukrania, antigua URSS) sufrió una explosión seguida por un incendio. La destrucción del reactor produjo un enorme escape de radioelementos que, formando una nube radiactiva, se fue desplazando impulsado por el viento y diseminó 131I, 137Cs, 90Sr y otras sustancias peligrosas a través de más de 200.000 km2 de todo el Este y Centro de Europa.
Los depósitos radiactivos más importantes se produjeron sobre unos 2600 km2 de superficie en una radio de 30 km alrededor de la central nuclear, superficie que fue rápidamente declarada “zona de exclusión” y de la que fueron inmediatamente evacuadas 116.000 personas (que nunca podrán regresar a sus casas). No obstante, hoy se estima que el territorio que quedó gravemente contaminado fue mucho más extenso y podría acercarse 28.000 km2 (en cuyo caso el número de personas que sufrieron una contaminación grave (>185 kBq/m² de 137Cs) podría ser de 830.000. Varios millones de personas recibieron radiación por encima de la norma y siguen viviendo en áreas con un nivel significativo de contaminación.
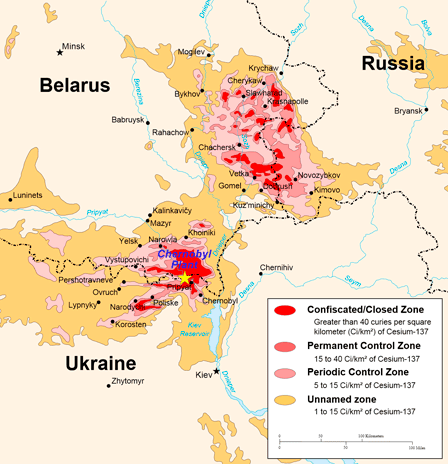 |
|
Áreas más afectadas por la contaminación radioactiva que produjo el accidente de Chernobil. Fuente: documento de dominio público disponible en: http://upload.wikimedia.org/wikipedia/commons/2/23/Chernobyl_radiation_map_1996.svg |
{kind=link}
Desde el punto de vista de la contaminación generada, la de Chernobil ha sido la mayor explosión nuclear de la historia ya que dispersó 400 veces más radiactividad que la de Hiroshima.
Las explosiones de bombas atómicas liberan radionucleidos con un periodo muy corto y que, por tanto, desaparecen muy deprisa del entorno. Gracias a ello en los lugares del mundo en los que se produjeron esas explosiones entre los años 40 y 70 la radiactividad es hoy residual y las condiciones ambientales muy próximas a las preexistentes. Sin embargo, en Chernobil y en sus alrededores (incluyendo superficies que hoy corresponden a Ucrania, Rusia y Bielorrusia) los acontecimientos han sido distintos y tras la destrucción masiva se ha producido una recolonización vegetal aunque persista una elevadísima tasa de radiactividad (que se va a mantener durante decenas de miles de años).
A lo largo del primer mes después de la explosión la radiactividad produjo la muerte de la mayoría de los organismos presentes en un radio de una decena de kilómetros. Los pinos (P.sylvestris) que formaban grandes masas boscosas alrededor de la instalación adquirieron un extraño color rojo ladrillo durante algunos días y después, prácticamente al unísono, murieron. Sus restos fueron arrancados con buldócer, quemados y enterrados in situ y, desde entonces, los pinos no han sido capaces de regenerarse (aunque el espacio liberado por la desaparición de sus bosques ha sido ocupado con posterioridad por abedulares o bosques de otras especies más resistentes a la radiactividad).
A mayor distancia los efectos variaron más dependiendo de las especies y mientras que las coníferas se mostraron muy frágiles, los caducifolios, herbáceas y musgos resistieron mucho mejor la radiactividad. La causa de esta desigual resistencia no se conoce aunque podría relacionarse con la distinta “anatomía” de los genomas ya que los problemas aparecen por la irradiación del ADN y cuanto más pequeños y compactos son los cromosomas menos probable es que resulten irradiados.
Pasados unos diez meses la mayoría de los isótopos habían dejado de irradiar y la radiactividad había bajado al 1% de la que llegó a registrarse en las primeras semanas. Gracias a ello, y aunque esta tasa es aún muy alta y resulta incompatible con una presencia humana prolongada, las plantas y animales más resistentes pudieron iniciar la recolonización del territorio a partir de la primavera de 1987.
Aunque las observaciones son aún insuficientes y algunas de las afirmaciones que se han hecho carecen del rigor científico necesario, todo parece indicar que algunos organismos experimentaron cambios fisiológicos o bioquímicos inducidos por la radiactividad. Por ejemplo, en las zonas más contaminadas el ADN de los pinos aparece muchas veces enrollado sobre sí mismo lo que reduce su vulnerabilidad frente a la radiación pero repercute en la morfología y metabolismo de los árboles que se vuelven más pequeños y crecen de forma más anárquica.
 |
|
Central de Chernobil vista desde una terraza de la ciudad abandonada de Pripiat. La vegetación está invadiendo las calles y edificios. FUENTE: Imagen de dominio público disponible en: http://es.wikipedia.org/wiki/Archivo:View_of_Chernobyl_taken_from_Pripyat.JPG. |
{kind=link}
Del mismo modo se han citado frecuentes casos de gigantismo u otras anomalías de origen genético en las plantas situadas en la zona de exclusión. De este modo, algunos abetos (Picea abies) estarían produciendo agujas de 4 ó 4,5 cm de longitud, dos veces más largas de lo que es habitual en esa especie, mientras que en los pinos (P. sylvestris) éstas alcanzarían 12 a 14 cm cuando lo habitual es que no superen 5. Por otra parte, en 1992 el número de agujas que producían los brotes jóvenes era el triple que en 1986 antes de la catástrofe y mayor el porcentaje de semillas que germinaban.
Todos estos hechos, que deberán documentarse mejor durante los próximos años pese a ser perfectamente verosímiles, indican claramente un aumento de la capacidad reproductiva y de crecimiento de las plantas que no puede haber sido consecuencia más que de la propia radiactividad.
Las observaciones que se han hecho en la fauna demuestran la existencia de impactos muy importantes pero hasta el momento su interpretación plantea más interrogantes que nuevas certezas.
La mayoría de los animales presentes en la zona de exclusión murió o sufrió un grave deterioro orgánico y fue incapaz de reproducirse tras quedar expuesta a la radiactividad. Esta fue la suerte del ganado que quedó abandonado en una isla a seis kilómetros del reactor tras la evacuación de la zona. Los caballos murieron rápidamente como consecuencia de la destrucción de sus glándulas tiroides y las pocas reses que sobrevivieron en la zona presentaron importantes disfunciones que supusieron su debilitamiento y rápida extinción. Sin embargo, con posterioridad los caballos han sido reintroducidos y grupos de individuos semisalvajes procedentes de las áreas menos expuestas se han ido instalando en el área de manera más o menos espontánea. Sorprendentemente, todas esas poblaciones se han acomodado sin problemas al entorno radiactivo y no parecen acusar sus consecuencias.
Algo parecido ocurrió con los pequeños roedores (Apodemus, Microtus) que desaparecieron durante la fase de radiación más intensa. Sin embargo, el vacío que dejaron fue ocupado en los años siguientes por otros individuos que llegaron de las regiones vecinas sin que se sepa por qué estas poblaciones de origen externo resisten mejor la radiación que la primera.
 |
|
Aspecto actual de los bosques en la “zona roja”. Los pinos murieron y sus restos fueron quemados y enterrados pero su lugar ha sido ocupado por abedules, mucho más tolerantes a la radiactividad. Fuente: imagen de dominio público diponible en http://www.flickr.com/photos/lord_yo/3758832940/sizes/l/in/photostream/. |
En las aves el número de alteraciones observadas es mayor en las especies ponedoras de grandes huevos y con plumajes vistosos e inferior en las más discretas. Entre las más afectadas se encuentran las currucas (Sylvia spp) y las golondrinas (Hirundo rustica) entre las que se han observado plumajes y huevos anormales y un menor éxito reproductivo.
El caso de los hongos es aún más sorprendente por la excepcional capacidad demostrada por algunos de ellos para sobrevivir en un entorno letal. Así, los investigadores han observado mediante robots la existencia de hongos melánicos creciendo sobre los restos de los muros del propio reactor y se ha demostrado que algunos taxones, como Cryptococcus neoformans o Cladosporium prosperan mejor en ambientes radiactivos gracias a la melanina que les permite aprovechar le energía de las radiaciones ionizantes del reactor.
Aunque la información disponible es aún escasa y la perspectiva histórica insuficiente, no hay duda de que la contaminación nuclear ha supuesto cambios muy importantes en la biodiversidad y estructura de los ecosistemas:
- Los pinares han sido sustituidos por praderas “naturales” o por abedulares.
- Numerosos grandes mamíferos que habían desaparecido a consecuencia de la presión humana han retornado a la zona aprovechando la tranquilidad que hoy encuentran en ella (lobo, lince, jabalí…).
- Varias especies emblemáticas de la fauna europea más amenazada han sido introducidas con éxito en el área para facilitar el aumento de sus poblaciones (caballo de Prjevalski, bisonte europeo).
- Algunas plantas cultivadas y animales domésticos que quedaron abandonados a su suerte tras la huida forzada de sus propietarios sobreviven y han recuperado rasgos o comportamientos propios de las especies salvajes de las que proceden. Es el caso, por ejemplo, de los perros que cazan formando manadas similares a las de los lobos.
Muchos animales son hoy más abundantes en la zona de exclusión que fuera de ella y la diversidad de plantas existente en las áreas más radiactivas es comparable a la de los espacios naturales protegidos de las regiones circundantes.
Si fuera preciso establecer un balance de las consecuencias de la catástrofe a la luz de lo que hoy se sabe, habría que confrontar dos tipos de hechos más o menos contradictorios:
- La radiactividad ha causado la muerte de la mayor parte de los organismos en un primer momento y, ulteriormente, ha originado impactos negativos muy significativos para la vida de numerosos animales y plantas. La recuperación que ha seguido al periodo inicial de mortandad ha sido muy desigual dependiendo de las especies por lo que los ecosistemas han quedado gravemente alterados.
- El accidente ha obligado a evacuar la zona y todas las actividades humanas han desaparecido de forma repentina e irreversible. Ello ha supuesto el desvanecimiento de la presión antrópica sobre el medio, ha permitido el retorno de numerosas especies extintas o que se encontraban en situación crítica y hace posible que los ecosistemas vuelvan a regirse por sus propias leyes naturales. La biodiversidad y la biomasa presentes en la zona han alcanzado así valores desconocidos desde hace mucho tiempo gracias a la desaparición de las interferencias humanas.
 |
|
Pasado el primer momento, las coníferas empiezan a regenerarse en algunas zonas de las que habían desaparecido. Algunos individuos presentan anomalías congénitas que podrían defenderles de las altas tasas de radiactividad. Foto: imagen de dominio público diponible en: |
{kind=link}
Lo acaecido en Chernobil ilustra muy bien una paradoja que tendemos a olvidar con demasiada frecuencia: un acontecimiento catastrófico para la humanidad puede no serlo (o no ser tan importante) para el medio natural. Al ser las actividades humanas “normales” más devastadoras a largo plazo para la biodiversidad que el mayor de los desastres industriales, la desaparición de la actividad humana tras uno de estos acontecimientos puede contribuir a una cierta recuperación de la naturaleza o, al menos, al restablecimiento de un nuevo equilibrio más próximo al natural que el que queda destruido por el desastre.
No obstante, quedan demasiadas cuestiones por resolver antes de poder establecer un balance definitivo y la falta de estudios a largo plazo sobre genética, demografía, tasas de mutación, esperanza de vida, fertilidad, salud u otros aspectos impide saber si las poblaciones expuestas de manera crónica a una elevada tasa de radiación difieren de las no expuestas y en qué medida ello es importante o no para su supervivencia. Mientras estas cuestiones no queden aclaradas no será posible conocer en su totalidad el efecto del desastre de Chernobil (o de otros similares ya ocurridos, como el de Fukushima, o por venir).